Мембранная ткань: воду не пропускает, дышать позволяет. Состав мембрана
Мембранная водонепроницаемая ткань: свойства, характеристики непромокаемости
Развитие технологий производства тканей позволило получить новое поколение многофункциональных материалов. Одним из них является мембрана — полупроницаемая плёнка с особой структурой. Мембранная ткань — это многослойные полотно, включающее такую плёнку. В готовых изделиях умные водоотталкивающие ткани не пропускают воду снаружи, но позволяют испаряться влаге, которая образуется внутри. Нижний слой мягкий, верхний — защитный, износостойкий. Средний — это защитный материал и мембрана.

Первая мембрана была произведена в 1969 году Уилбертом Гором и его сыном Робертом (Wilbert L. Gore и Robert W. Gore). Она была предназначена для использования в космосе, название было запатентовано как Gore-Tex (гортэкс). Производится она из фторопласта (тефлона). После истечения срока действия патента появились другие похожие водонепроницаемые материалы для одежды, которые используются для производства спецодежды и обуви. Например, синтетическая ткань «алова», которая состоит из 100 % трикотажного полиэстера снаружи и мембраны с внутренней стороны.
Преимущества
 Основными свойствами ткани являются:
Основными свойствами ткани являются:
- водонепроницаемость. Выражается в числовом эквиваленте. Он обозначает давление воды, которое выдержит ткань. Существуют различные показатели, на которые следует обращать внимание при покупке таких изделий: цифра 3.000 значит, что ткань способна выдержать мелкий дождь и несильный снег, 10.000 — сильный дождь, 20.000 — ткань не промокнет в сильную непогоду и в штормовых условиях;
- паровыводимость. Также выражается в числовом эквиваленте — количество пара в граммах на квадратный метр ткани, который она выводит за сутки. Чем выше показатель, тем ткань лучше;
- защита от ветра.
Виды
Производится мембрана из органических и неорганических материалов. Существует несколько типов:- поровая (тефлоновая). Имеет микропоры на поверхности внешнего слоя, которые не пропускают воду, но позволяют свободно испаряться влаге (диффузия молекул), собирающейся внутри. Недостатком является то, что поры могут забиваться, и тогда система испарения нарушается;
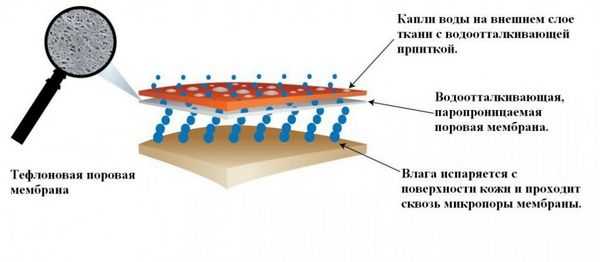
- беспоровая (полиуретановая). Не имеет пор на поверхности, не пропускает воду. Влага, которая образуется внутри изделия, сначала скапливается на внутренней поверхности внешнего слоя, затем постепенно испаряется. Недостатком можно считать то, что влага испаряется не сразу, и может появляться ощущение, что изделие мокрое;

полиуретановая мембрана
- комбинированная. Внутри такого материала находится поровая мембрана, а сверху неё ещё один защитный слой, который защищает поры от забивания. Этот вид ткани сочетает в себе преимущества двух первых.
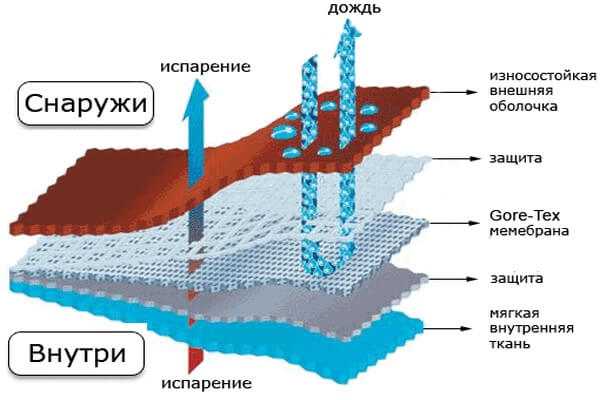
Строение
По структуре мембранные ткани делятся на:
- двухслойные. В такой ткани соединены мембрана и внешняя поверхность, подкладка не закреплена. Она хорошо «дышит», гибкая и относительно недорогая;
- трёхслойные. Верхний слой склеен с мембраной и подкладкой (сеткой). Такой материал получается более лёгким, меньше пропускает воздух, его называют ламинированной тканью. Цена на него самая высокая;
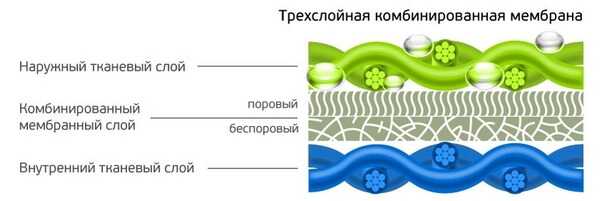
- мембрана в 2,5 слоя. Вместо подкладки или сетки используется вспененное пупырчатое нанесение, защищающее мембрану.
Применение
Сейчас этот материал используют для производства одежды и обуви для людей, ведущих активных образ жизни. Из неё шьют куртки, брюки, комбинезоны, спортивные костюмы и обувь. Это незаменимый материал для альпинистов, спортсменов, туристов, людей, занимающихся экстремальными видами спорта. В последние годы из него шьют и детскую верхнюю одежду.

Так как сама мембрана не защищает от холода, одежда из неё бывает демисезонной и с утеплителем (флис).Куртки и комбинезоны из мембранной ткани могут окрашиваться в самые разнообразные оттенки, большой популярностью пользуются и камуфляжные расцветки.
Польза
 Помимо отличных гигиенических и защитных свойств, одежда с мембраной ценится за:
Помимо отличных гигиенических и защитных свойств, одежда с мембраной ценится за:
- лёгкость;
- прочность;
- удобство;
- яркие расцветки.
Как носить
Основным правилом ношения одежды с мембраной является многослойность. Нужно надевать нижний слой (бельё), средний (свитер) и мембрану. При этом, лучше, если внутренняя одежда будет содержать определённый объём синтетики, чтобы пропускать через себя испарения.
Как ухаживать
 Мембранную одежду нельзя стирать обычными порошками, так как они могут повредить структуру, забить поры.
Мембранную одежду нельзя стирать обычными порошками, так как они могут повредить структуру, забить поры.
- Нужно использовать специальные средства для стирки, хозяйственное или жидкое мыло. Отжимать в стиральной машине запрещается. При ручной стирке нельзя сильно скручивать изделие. Если имеются сильные загрязнения, их можно удалить при помощи мягкой щётки.
- После стирки воде нужно дать стечь, затем повесить вещь на открытом воздухе или в комнате вдали от отопительных приборов и не допускать попадания солнечных лучей.
- При производстве одежды на её поверхность наносится специальное покрытие DWR (Durable Water Repellence), которое обеспечивает ей дополнительную защиту от влаги. После многократных стирок это покрытие исчезает, поэтому рекомендуется восстанавливать его каждый раз после стирки при помощи специального спрея. Распыляется такое покрытие именно на сухую чистую поверхность.
- Гладить мембранную одежду нельзя, так при контакте с нагретым утюгом повредится её структура.
Обувь
Мембрана располагается в обуви в виде носка, обычно не доходит до самого верха. Её расположение также зависит и от самой конструкции обуви, наличия молнии или языка. В ботинке создаётся особый микроклимат, испарения от человеческого тела проходят через мембрану и выходят наружу благодаря разнице внутреннего и внешнего давления. Сверху обувь обычно защищена водонепроницаемой и износостойкой тканью.
На заметку
Так же как и с верхней одеждой, здесь работает принцип многослойности — под мембранную обувь необходимо надевать носки. Они должны быть не 100 % хлопковые или шерстяные, а содержать минимум 10% синтетики. Тогда пар будет отводиться.
Если вода попадёт через верхний слой, то мембрана его не пропустит, но вода останется в обуви. Поэтому очень важно хорошо её просушивать.
Уход за обувью
- Нельзя допускать налипания грязи на поверхность, так как поры должны быть открытыми.
- Чистить мембранную обувь нужно сухой щёткой или губкой, смоченной в мыльной воде.
- Сушить вдали от батарей и других источников тепла, можно положить внутрь газету.
- Каждый раз после чистки обуви желательно обрабатывать её водоотталкивающими спреями.
Промышленность
 При производстве различных машинных приборов, насосов, карбюраторов и т.д. применяют прорезиненную мембрану. Это техническая ткань, вулканизированная с двух сторон.
При производстве различных машинных приборов, насосов, карбюраторов и т.д. применяют прорезиненную мембрану. Это техническая ткань, вулканизированная с двух сторон.
Из синтетической аловы шьют защитные чехлы и обивку для мебели.
Высокотехнологичные мембраны для одежды рушат все старые представления о тёплом непромокаемом изделии: оно не тяжелое, в нем комфортно и взрослым, и детям, обладает отличными эстетическими качествами. А при правильной эксплуатации и надлежащем уходе изделия из мембранной ткани прослужат очень долго.
© 2018 textiletrend.ru
textiletrend.ru
| классифицируют в соответствии с размером углеводной части, которая может быть представлена всего лишь одним моносахаридным остатком, с одной стороны, и очень сложным углеводным полимером — с другой. Моногликозилцерамиды обычно называют цереброзидами. Ганглиозиды представляют собой класс анионных гликосфинголипидов, которые содержат один или несколько остатков сиаловой кислоты, связанных с сахарными остатками церамидолигосахарида. Глобозидами называют нейтральные гликосфинголипиды, которые не содержат остатков отрицательно заряженной сиаловой кислоты. Гликосфинголипиды находятся на наружной поверхности плазматических мембран животных клеток; обычно они являются минорными компонентами, но иногда содержатся в значительных количествах. Моногалактозилцерамид — это один из основных компонентов миелиновой оболочки нервного волокна. В некоторых случаях гликосфинголипиды локализуются не в плазматической мембране, а во внутриклеточных мембранах. Гликосфинголипиды мембран эритроцитов несут антигены группы крови. В клетках аденокарциномы человека накапливаются необычные фукозилированные гликосфинголипиды, которые можно использовать для обнаружения этих клеток и контроля за развитием опухоли. Стеролы Эти липиды присутствуют во многих мембранах растений, животных и микробов. По-видимому, самым распространенным из сте-ролов является холестерол. Его молекула состоит из компактного, жесткого гидрофобного ядра, а полярной головкой является гидро-ксильная группа. Холестерол содержится в плазматических мембранах животных клеток, в лизосомах, эндосомах и в мембранах аппарата Гольджи. Он составляет около 30% всей массы мембранных липидов во многих плазматических мембранах животных клеток. В высших растениях обнаружены другие стеролы, чаще всего ситостерол и стигмастерол. Растительные стеролы часто имеют еще одну боковую цепь в положении С-24 и/или двойную связь в положении С-22. В мембранах дрожжей и дру-
гих эукариотических микроорганизмов часто содержится эргостерол. К классу стеролоподобных липидов относят также гопанои-ды, которые найдены в бактериях и некоторых растениях. Минорные компоненты В мембранах присутствуют также и другие липиды, которые можно отнести к разряду минорных компонентов вследствие их малого содержания в мембранах. Так, в мембранах обычно обнаруживаются, хотя и в очень малых количествах, свободные жирные кислоты и лизофосфолипиды. Пожалуй, исключением из этого правила являются мембраны хромаффинных гранул, которые, как известно, содержат необычно много свободных жирных кислот. Минорными компонентами мембран являются также моноацил- и диацилглице-ролы. Диацилглицеролы выполняют важную функцию вторых посредников в передаче сигнала при активации клеток рядом биологически активных веществ. Эта система клеточного отклика на внешний стимул будет детально рассмотрена в гл. 9. В мембранах обычно присутствуют и полиизопреноидные липиды. К ним относятся уби-хиноны и менахиноны — компоненты цепи электронного транспорта в мембранах. Можно отметить также ундекапренол и долихол, которые являются липидными переносчиками промежуточных продуктов соответственно при биосинтезе клеточной стенки у прокариот и при биосинтезе гликопротеинов в аппарате Гольджи эукариот. Длина молекул этих липидов в вытянутом состоянии значительно превышает толщину бислоя, поэтому неизвестно, как эти молекулы в нем расположены. Неясно также, почему липидными переносчиками служат именно полиизопреноидные структуры. 2. МНОГООБРАЗИЕ ФУНКЦИЙ МЕМБРАННЫХ ЛИПИДОВ Совершенно очевидно, что липидный состав различных мембран не является случайным, однако удовлетворительного объяснения этому феномену не найдено. Любая конкретная мембрана может содержать более ста разных типов липидных молекул. Почему их так много и почему каждая мембрана имеет уникальный липидный состав? Пути биосинтеза мембранных липидов и механизмы их доставки к местам назначения обсуждаются в разд. 10.4. Становится все более очевидным, что липиды активно участвуют в процессах, протекающих в мембранах, однако причины их разнообразия неясны. Рассмотрим некоторые факторы, возможно, определяющие липидный состав мембраны. 1. Смесь липидов обязательно должна быть способна образовать стабильный бислой, в котором могли бы функционировать белки.
Как было показано экспериментально, организмы часто могут выдерживать — причем без всяких последствий — существенные изменения липидного состава мембран. Например, с помощью генетической трансформации можно получить штаммы Е. coli, в мембранах которых содержится 34% фосфатидной кислоты, обычно отсутствующей в штаммах дикого типа. Очевидно, тот липидный состав, который характерен для штаммов дикого типа, не является обязательным для выживания клеток, по крайней мере, в условиях их выращивания в лаборатории. 3. МЕМБРАННЫЕ БЕЛКИ Как видно из табл. 1.3 и 1.4, мембраны содержат от 20 до 80% белка. Как правило, именно белки ответственны за функциональную активность мембран. К ним относятся разнообразные ферменты, транспортные белки, рецепторы, каналы, поры и т. д., которые обеспечивают уникальность функций каждой мембраны. Первые успехи в изучении мембранных белков были достигнуты тогда, когда биохимики научились использовать детергенты для выделения мембранных белков в функционально активной форме. Это были работы по изучению ферментных комплексов внутренней мембраны митохондрий. Значительным шагом вперед было осознание того, что мембранные белки имеют не исключительно /3-складчатую структуру, как предполагалось в модели «элементарной мембраны» Дэвсона—Даниелли—Робертсона, а содержат достаточно много а-спиралей. Важное значение имел также вывод о том, что мембранные белки могут глубоко проникать в липидный бислой или даже пронизывать его и что их стабилизация осуществляется за счет гидрофобных взаимодействий. Эти термодинамические представления существенно обогатили принцип «гидрофобных сил», предложенный для объяснения структуры белков и предполагавший существование неполярной, гидрофобной области внутри белковой глобулы и полярных, гидрофильных участков, контактирующих с водной средой. По мере совершенствования методов очистки удавалось получать в изолированном виде все большее число мембранных белков. Определение первичной структуры большинства из них было затруднено из-за плохой растворимости в воде как самих белков, так и получаемых из них гидрофобных пептидов. В середине 1970-х гг. эта проблема была решена для двух мембранных белков — гликофорина и цитохрома bs, что позволило установить основной принцип структурной организации интегральных белков. В аминокислотной последовательности гликофорина — сиалогликопротеина из мембраны эритроцитов — был обнаружен короткий участок, состоящий из 23 неполярных аминокислот и расположенный примерно в середине цепи. Данные топологических и других исследований показали, что молекула гликофорина полностью пронизывает мембрану, причем погруженный в мембрану гидрофобный участок имеет а-спиральную конфигурацию. Так вошла в жизнь новая, теперь уже общепризнанная концепция о наличии в мембранных белках а-спиральных доменов, пронизывающих мембрану. Эта концепция была полностью подтверждена при изучении трансмембранных белков с помощью методов, которые позволяют получить максимально возможное в наше время разрешение. Судя по результатам реконструкции электронно-микроскопических изображений препаратов бактериородопсина из пурпурной мембраны Halobacterium halobium и по данным рентгеноструктурного исследования фотосинтетических реакционных центров бактерий, эти белки содержат несколько а-спиральных участков, последовательно пересекающих бнслой. Другой вариант расположения полипептидной цепи в мембране был обнаружен при изучении аминокислотной последовательности интактной формы микросомного цитохрома bi. Было показано, что этот белок содержит относительно короткий участок вблизи карбоксильного конца, состоящий из гидрофобных аминокислот. Этот «гидрофобный якорь» можно было удалить с помощью протеолиза, причем гемсвязывающий домен высвобождался в водорастворимой форме. Локализованный в мембране гидрофобный домен, или «якорь», стал еще одним характерным элементом структуры мембранных белков. В основе современных представлений о структуре мембранных белков лежит идея о том, что их полипептидная цепь уложена так, чтобы образовалась неполярная, гидрофобная поверхность, контактирующая с неполярной областью липидного бислоя. Полярные или заряженные домены белковой молекулы могут вазимодействовать с полярными головками липидов на поверхности бислоя. Многие мембранные белки являются трансмембранными и пронизывают бислой. Некоторые белки, по-видимому, связаны с мембраной лишь за счет их взаимодействия с другими белками. Мембранные белки обычно связываются с мембраной с помощью нековалентных взаимодействий — гидрофобных или электростатических сил. Однако есть мембранные белки, которые связаны с липидами ковалентно. Такие примеры пока немногочисленны, но их появляется все больше. Многие белки плазматических мембран растительных и животных клеток относятся к классу гликопротеинов. Углеводные остатки этих белков всегда находятся с наружной стороны плазматической мембраны. Обычно мембранные белки подразделяют на наружные и внутренние. При этом критерием служит степень жесткости обработки, необходимой для извлечения этих белков из мембраны. Периферические белки высвобождаются при промывании мембран буферными растворами с низкой ионной силой, буферными растворами с низким или, наоборот, высоким значением рН и в присутствии хелатирующих агентов, связывающих двухвалентные катионы. Как полагают, такие белки связаны с поверхностью мембраны за счет слабых электростатических взаимодействий с полярными головками липидных молекул либо с молекулами других белков. Часто бывает нелегко отличить периферические мембранные белки от белков, связавшихся с мембраной в процессе ее выделения. При обработке мембранного препарата буфером с низкой ионной силой в раствор переходит около 30% белков, связанных с мембраной эритроцитов. При несколько более жесткой обработке хаотропными агентами высвобождаются периферические белки. В ряде случаев эти агенты оказывают влияние достаточно сильное, чтобы разрушить белок-белковые взаимодействия, хотя денатурации белков при этом не происходит. Действие хаотропных агентов, «нарушающих структуру воды», обусловлено главным образом ослаблением гидрофобных взаимодействий между компонентами мембраны. Для высвобождения интегральных мембранных белков необходимо использовать детергенты или даже органические растворители. Детергенты разрушают липидный бислой и, как полагают, связываются с гидрофобными участками мембранных белков, контактирующими с гидрофобной областью бислоя. Для того чтобы сохранить интегральные мембранные белки в растворенном монодисперсном состоянии, в растворе постоянно должны присутствовать детергенты. При удалении детергентов неизбежно происходят агрегация белков и их последующее осаждение. Дальнейшая информация о взаимодействиях между белками и детергентами, а также о структуре и свойствах мембранных белков содержится в гл. 3. Резюме Методы дифракции рентгеновских лучей и электронной микроскопии сыграли историческую роль в развитии мембранологии и внесли решающий вклад в современные представления о биологических мембранах. Сегодня не вызывает сомнений, что липидный бислой образует структурную основу практически всех биологических мембран и что их функциональное многообразие основано именно на этом структурном единстве различных мембранных систем. Каждая отдельно взятая мембрана содержит большое число различающихся по своим химическим свойствам липидов. Причины этого разнообразия неясны, хотя появляется все больше данных об уникальных биологических функциях отдельных липидов. На долю белков приходится от 20 до 80% массы мембран. Многие из этих белков полностью пронизывают липидный бислой, и их удается солюбилизировать только с помощью детергентов. Другие мембранные белки, называемые периферическими, легкое извлекаются из мембраны с помощью буферных растворов с изменяющимися рН или ионной силой либо при удалении двухвалентных катионов с помощью хелатирующих агентов. В настоящее время разработаны методы выделения и характеристики индивидуальных мембран из клеток прокариот, из животных и в меньшей степени из растительных клеток. Разделение чаще всего основано на различиях в размере и плотности мембранных частиц, содержащихся в гомогенате разрушенных клеток. Можно также использоать различия в поверхностных свойствах мембран или в их электрофоретическом поведении. Выделение и очистка мембран — первый и обязательный этап в их биохимическом исследовании. |
www.coolreferat.com
Состав мембран - часть 2
гих эукариотических микроорганизмов часто содержится эргостерол. К классу стеролоподобных липидов относят также гопанои-ды, которые найдены в бактериях и некоторых растениях.
Минорные компоненты
В мембранах присутствуют также и другие липиды, которые можно отнести к разряду минорных компонентов вследствие их малого содержания в мембранах. Так, в мембранах обычно обнаруживаются, хотя и в очень малых количествах, свободные жирные кислоты и лизофосфолипиды. Пожалуй, исключением из этого правила являются мембраны хромаффинных гранул, которые, как известно, содержат необычно много свободных жирных кислот. Минорными компонентами мембран являются также моноацил- и диацилглице-ролы. Диацилглицеролы выполняют важную функцию вторых посредников в передаче сигнала при активации клеток рядом биологически активных веществ. Эта система клеточного отклика на внешний стимул будет детально рассмотрена в гл. 9. В мембранах обычно присутствуют и полиизопреноидные липиды. К ним относятся уби-хиноны и менахиноны — компоненты цепи электронного транспорта в мембранах. Можно отметить также ундекапренол и долихол, которые являются липидными переносчиками промежуточных продуктов соответственно при биосинтезе клеточной стенки у прокариот и при биосинтезе гликопротеинов в аппарате Гольджи эукариот. Длина молекул этих липидов в вытянутом состоянии значительно превышает толщину бислоя, поэтому неизвестно, как эти молекулы в нем расположены. Неясно также, почему липидными переносчиками служат именно полиизопреноидные структуры.
2. МНОГООБРАЗИЕ ФУНКЦИЙ МЕМБРАННЫХ ЛИПИДОВ
Совершенно очевидно, что липидный состав различных мембран не является случайным, однако удовлетворительного объяснения этому феномену не найдено. Любая конкретная мембрана может содержать более ста разных типов липидных молекул. Почему их так много и почему каждая мембрана имеет уникальный липидный состав? Пути биосинтеза мембранных липидов и механизмы их доставки к местам назначения обсуждаются в разд. 10.4. Становится все более очевидным, что липиды активно участвуют в процессах, протекающих в мембранах, однако причины их разнообразия неясны. Рассмотрим некоторые факторы, возможно, определяющие липидный состав мембраны.
1. Смесь липидов обязательно должна быть способна образовать стабильный бислой, в котором могли бы функционировать белки.
2.Некоторые липиды способствуют стабилизации сильно искривленных участков мембраны, образованию контакта между мембранами или связыванию определенных белков, поскольку форма этих молекул благоприятствует нужной упаковке бислоя на соответствующих участках мембраны.
3.Некоторые липиды являются важными биорегуляторами. Наиболее изучена в этом отношении регуляторная роль производных фосфатидилинозитола в плазматических мембранах клеток эукариот.
4.Некоторые липиды участвуют в реакциях биосинтеза. Например, в клетках Е. coliфосфатидилглицерол поставляет глицерофосфатный фрагмент при биосинтезе периплазматических олигосахари-дов.
5.Отдельные липиды необходимы для поддержания оптимальной активности ряда ферментов. Этот вопрос рассматривается в гл. 6.
6.Ганглиозиды, как полагают, играют важную роль в регуляции роста клеток, являются специфическими рецепторами в плазматической мембране и ответственны за клеточную адгезию.
Как было показано экспериментально, организмы часто могут выдерживать — причем без всяких последствий — существенные изменения липидного состава мембран. Например, с помощью генетической трансформации можно получить штаммы Е. coli, в мембранах которых содержится 34% фосфатидной кислоты, обычно отсутствующей в штаммах дикого типа. Очевидно, тот липидный состав, который характерен для штаммов дикого типа, не является обязательным для выживания клеток, по крайней мере, в условиях их выращивания в лаборатории.
3. МЕМБРАННЫЕ БЕЛКИ
Как видно из табл. 1.3 и 1.4, мембраны содержат от 20 до 80% белка. Как правило, именно белки ответственны за функциональную активность мембран. К ним относятся разнообразные ферменты, транспортные белки, рецепторы, каналы, поры и т. д., которые обеспечивают уникальность функций каждой мембраны. Первые успехи в изучении мембранных белков были достигнуты тогда, когда биохимики научились использовать детергенты для выделения мембранных белков в функционально активной форме. Это были работы по изучению ферментных комплексов внутренней мембраны митохондрий. Значительным шагом вперед было осознание того, что мембранные белки имеют не исключительно /3-складчатую структуру, как предполагалось в модели «элементарной мембраны» Дэвсона—Даниелли—Робертсона, а содержат достаточно много а-спиралей. Важное значение имел также вывод о том, что мембранные белки могут глубоко проникать в липидный бислой или даже пронизывать его и что их стабилизация осуществляется за счет гидрофобных взаимодействий. Эти термодинамические представления существенно обогатили принцип «гидрофобных сил», предложенный для объяснения структуры белков и предполагавший существование неполярной, гидрофобной области внутри белковой глобулы и полярных, гидрофильных участков, контактирующих с водной средой.
По мере совершенствования методов очистки удавалось получать в изолированном виде все большее число мембранных белков. Определение первичной структуры большинства из них было затруднено из-за плохой растворимости в воде как самих белков, так и получаемых из них гидрофобных пептидов. В середине 1970-х гг. эта проблема была решена для двух мембранных белков — гликофорина и цитохрома bs, что позволило установить основной принцип структурной организации интегральных белков. В аминокислотной последовательности гликофорина — сиалогликопротеина из мембраны эритроцитов — был обнаружен короткий участок, состоящий из 23 неполярных аминокислот и расположенный примерно в середине цепи. Данные топологических и других исследований показали, что молекула гликофорина полностью пронизывает мембрану, причем погруженный в мембрану гидрофобный участок имеет а-спиральную конфигурацию. Так вошла в жизнь новая, теперь уже общепризнанная концепция о наличии в мембранных белках а-спиральных доменов, пронизывающих мембрану. Эта концепция была полностью подтверждена при изучении трансмембранных белков с помощью методов, которые позволяют получить максимально возможное в наше время разрешение. Судя по результатам реконструкции электронно-микроскопических изображений препаратов бактериородопсина из пурпурной мембраны Halobacteriumhalobiumи по данным рентгеноструктурного исследования фотосинтетических реакционных центров бактерий, эти белки содержат несколько а-спиральных участков, последовательно пересекающих бнслой.
Другой вариант расположения полипептидной цепи в мембране был обнаружен при изучении аминокислотной последовательности интактной формы микросомного цитохрома bi. Было показано, что этот белок содержит относительно короткий участок вблизи карбоксильного конца, состоящий из гидрофобных аминокислот. Этот «гидрофобный якорь» можно было удалить с помощью протеолиза, причем гемсвязывающий домен высвобождался в водорастворимой форме. Локализованный в мембране гидрофобный домен, или «якорь», стал еще одним характерным элементом структуры мембранных белков.
В основе современных представлений о структуре мембранных белков лежит идея о том, что их полипептидная цепь уложена так, чтобы образовалась неполярная, гидрофобная поверхность, контактирующая с неполярной областью липидного бислоя. Полярные или заряженные домены белковой молекулы могут вазимодействовать с полярными головками липидов на поверхности бислоя. Многие мембранные белки являются трансмембранными и пронизывают бислой. Некоторые белки, по-видимому, связаны с мембраной лишь за счет их взаимодействия с другими белками.
Мембранные белки обычно связываются с мембраной с помощью нековалентных взаимодействий — гидрофобных или электростатических сил. Однако есть мембранные белки, которые связаны с липидами ковалентно. Такие примеры пока немногочисленны, но их появляется все больше. Многие белки плазматических мембран растительных и животных клеток относятся к классу гликопротеинов. Углеводные остатки этих белков всегда находятся с наружной стороны плазматической мембраны.
Обычно мембранные белки подразделяют на наружные и внутренние. При этом критерием служит степень жесткости обработки, необходимой для извлечения этих белков из мембраны. Периферические белки высвобождаются при промывании мембран буферными растворами с низкой ионной силой, буферными растворами с низким или, наоборот, высоким значением рН и в присутствии хелатирующих агентов, связывающих двухвалентные катионы. Как полагают, такие белки связаны с поверхностью мембраны за счет слабых электростатических взаимодействий с полярными головками липидных молекул либо с молекулами других белков. Часто бывает нелегко отличить периферические мембранные белки от белков, связавшихся с мембраной в процессе ее выделения. При обработке мембранного препарата буфером с низкой ионной силой в раствор переходит около 30% белков, связанных с мембраной эритроцитов. При несколько более жесткой обработке хаотропными агентами высвобождаются периферические белки. В ряде случаев эти агенты оказывают влияние достаточно сильное, чтобы разрушить белок-белковые взаимодействия, хотя денатурации белков при этом не происходит. Действие хаотропных агентов, «нарушающих структуру воды», обусловлено главным образом ослаблением гидрофобных взаимодействий между компонентами мембраны.
mirznanii.com
Биофизика клеточных процессов. Мембраны, их состав, структура. Мембранный транспорт.
Биологическими мембранами называют функциональные структуры клеток, ограничивающих цитоплазму и большинство внутриклеточных структур. Мембраны также образуют внутри клетки единую систему канальцев и полостей. Толщина биологических мембран составляет 7-10 нм, однако вследствие большой общей площади мембран и их высокой плотности, их масса составляет более 50 % сухой массы клетки. Биологические мембраны являются высокоизбирательными барьерами проницаемости. Поток молекул в клетку и из клетки, в органоид и из нее, контролируется находящимися в мембранах специфическими системами транспорта. Транспортные процессы в мембране регулируют объем клетки и поддерживают ионный состав, необходимый для работы ферментативныхсистем, т.е. гомеостаз клетки. Транспортные процессы также создают ионные градиенты, необходимые для создания мембранного потенциала, поддержания возбудимости, а также транспорта некоторых молекул и ионов. Системы транспорта в мембране переносят из внешней среды и концентрируют в компартментах клетки вещества, необходимые для функционирования клетки. Так как клетки и внутриклеточные структуры, с точки зрения термодинамики, открытые системы, они постоянно обмениваются с окружающей средой и веществом и энергией. Поэтому транспорт вещества и энергии через мембраны - необходимое условие существования живых систем.
Состав и структура биомембран.
Состав мембран зависит от их типа и функций, однако во всех типах биологических мембран основными структурными компонентами являются молекулы липидов и белков. Некоторые мембраны содержат также и углеводы, которые связаны или с белками (гликопротеины) или с липидами (гликолипиды). Важным структурным компонентом мембран является вода. Взаимодействие молекул липидов, белков с водой определяет специфические структурно-функциональные свойства мембраны и определяет стабильность мембранных структур.
В состав биологических мембран входят молекулы, относящиеся к различным классам липидов, а также стероиды. Мембранные липиды имеют сравнительно небольшую полярную (заряженную) головку и длинные неполярные (незаряженные) углеводородные цепи. Жирные кислоты, входящие в состав липидов мембран, могут быть насыщенными и ненасыщенными двойными связями и обычно имеют 14-22 углеродных атома.
У фосфолипидов (фосфоглицеридов) неполярная часть представлена двумя остатками жирных кислот, этерефицирующих две гидроксильные группы глицерола. Третья гидроксильная группа образует полярную головку – остаток фосфорной кислоты. К остатку фосфорной кислоты может быть присоединен остаток аминоспирта (например, фосфатидилхолин), сахара (фосфатидилинозитол), аминокислоты (фосфатидилсерин).
Сфинголипиды построены из одного остатка жирной кислоты, из одного остатка длинноцепочечного аминоспирта сфингозина (или его производного). Полярная головка этих молекул представлена остатком спирта.
Из стероидов наиболее распространенным компонентом мембран является холестерин (холестерол) и его производные. Содержание холестерина характерно для мембран эукариотических клеток, у большинства прокариот он не обнаружен.
Белковый состав мембран также исключительно многообразен. Мембраны многих клеток содержат большое число белков, с молекулярной массой 10 до 250 кДа. Молекулы белков могут быть либо частично, либо целиком, погружены в липидный бислой или могут пронизывать его насквозь. Это так называемые интегральные белки. Белки, которые слабо удерживаются на мембране за счет слабых, в основном электростатических, взаимодействий, называются периферическими. Молекулы белков в составе мембран выполняют ферментативные, транспортные, регуляторные и опорно-строительную функции. Относительное содержание белков и липидов в составе мембран зависит от типа клеток и от специфики мембранных функций. Так, в составе внутриклеточных мембран (митохондрий, микросом) содержание белка выше, чем содержание липидов. В мембранах нервных клеток млекопитающих содержание белка составляет всего 20-25 % по сухой массе ( табл.1)
Таблица 1
Относительное содержание белков и липидов в составе различных мембран (% от сухой массы)
| Вид мембран | белки | липиды |
| Миелиновая оболочка нервных клеток млекопитающих | ||
| Плазмалемма эритроцитов человека | ||
| Плазмалемма клеток печени млекопитающих | ||
| Наружная мембрана митохондрий млекопитающих | ||
| Внутренняя мембрана митохондрий млекопитающих | ||
| Микросомы клеток печени млекопитающих |
Относительное содержание представителей того иного класса липидов, белковых молекул, также зависит от структурных особенностей и функциональных свойств мембраны. Так, мембрана нервных клеток, содержит небольшое количество белка ( 18 % массы мембраны). В плазматической мембране большинства клеток содержание белка достигает до 50 % массы, так как различные ионные насосы, каналы, рецепторы, ферменты представлены, в основном, молекулами белков. Наиболее высоким содержанием белков характеризуются мембраны внутренние митохондрий и хлоропластов, где располагаются электронно- транспортные цепи и происходит синтез АТФ. В составе мембран большинства типов клеток наиболее распространены следующие молекулы липидов: фосфатидилхолин (лецитин), фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозитол. Мембраны эукариотических клеток богаты также холестерином.
В таблице 2 приведены данные о липидном составе различных мембран клеток млекопитающих.
Таблица 2
Содержание липидов в составе плазматической мембраны и мембран органоидов животной клетки ( в % к общей массе липидов)
| Липиды | Плазмалемма | Митохондрии | Лизосомы | Ядро | ЭПС | Аппа рат Гольджи |
| Фосфатидил холин | ||||||
| Сфингомиелин | ||||||
| Фосфатидилэтаноламин | 11,5 | 28,5 | 12,5 | 16.5 | ||
| Фосфатидилсерин | 3,5 | 2,5 | ||||
| Фосфатидилинозитол | 2,5 | 7,5 | ||||
| Лизофосфатидилхолин | 2,5 | 1,5 | ||||
| Дифосфатидилглицерин | ||||||
| Холестерин и ее эфиры | ||||||
| Свободные жирные кислоты | - | - | ||||
| Другие липиды | 2,5 | 5,5 |
Воду, входящую в состав мембран, подразделяют на связанную, захваченную и свободную. Связанная вода - это вода гидратных оболочек ионов и полярных участков липидов и белков. Гидратные оболочки основных структурообразующих липидов состоят из 10-12 молекул воды. Эта вода осмотически и химически неактивна и не способна растворять какие - либо вещества. Захваченная вода находится в основном между двумя слоями в липидов мембраны. По подвижности, химической активности она занимает промежуточное положение между связанной и свободной водой. Свободная вода входит в состав мембраны в виде самостоятельной фазы и обладает всеми свойствами жидкой воды.
Похожие статьи:
poznayka.org
| митохондрии | микро-сомы | ли10- сомы | плазматическая мембрана | ядерная мембрана | мембраны аппарата Гольджи | |
| Кардиолипин | 18 | 1 | 1 | 1 | 4 | 1 |
| Фосфатидилэтаноламин | 35 | 22 | 14 | 23 | 13 | 20 |
| Фосфатидилхолин | 40 | 58 | 40 | 39 | 55 | 50 |
| Фосфатидилииозитол | 5 | 10 | 5 | 8 | 10 | 12 |
| Фосфатидилсерин | 1 | 2 | 2 | 9 | 3 | 6 |
| Фосфатидная кислота | — | 1 | 1 | 1 | ||
| Лиэофосфоглицериды 2' | 1 4 | 11 | 7 | 2 | 3 | 3 |
| Сфингомиелин | 1 | 1 | 20 | 16 | 3 | 8 |
| Фосфолипиды | 0,175 | 0,374 | 0,156 | 0,672 | 0,500 | 0,825 |
| Холестерол | 0,003 | 0,014 | 0,038 | 0,128 | 0,038 | 0,078 | Длина цепн: степень |
| ненасыщенности | ||||||
| Лаурииовая кислота | 12:0 | |||||
| Миристиновая кислота | 14:0 | |||||
| Пальмитиновая кислота | 16:0 | |||||
| Пальмитолеиновая кислота | 16:1 (9-цис) | |||||
| Стеариновая кислота | 18:0 | |||||
| Олеиновая кислота | 18:1 {9-цис) | |||||
| Вакценовая кислота | 18:1 (\\-цис) | |||||
| Линолевая кислота | 18:2 (9-цис, 12-цис) | |||||
| 7-Линоленовая кислота | 18:3 (6-цис, 9-цис, 12-цис) | |||||
| а-Линоленовая кислота | 18:3 (9-цис, \2-цис, 15-цис) | |||||
| Арахиновая кислота | 20:0 | |||||
| Бегеновая кислота | 22:0 | |||||
| Арахидоновая кислота | 20:4 (5-цис, %-цис, \\-цис, 14-цис) |

 Состав мембран
Состав мембранdoc4web.ru
Структура - мембрана - Большая Энциклопедия Нефти и Газа, статья, страница 1
Структура - мембрана
Cтраница 1
Структура мембраны как тилакоидов гран, так и тила-коидов стромы ( одиночных тилакоидов) была подробно изучена с помощью электронной микроскопии и иммунологических методов. Много информации получено также и о химическом составе этих мембран. [2]
Структура мембраны асимметрична, ее липидный матрикс состоит из двух слоев, причем наружный слой толще внутреннего. В липидный матрикс погружено множество белковых частиц, которые видны как на наружной, так и на внутренней поверхности тилакоида. Они видны также по обе стороны от гидрофильной центральной зоны внутри мембраны на срезе препарата, приготовленного методом замораживания - скалывания. Эти частицы могут совершенно свободно перемещаться в жидком липидном матриксе. Их содержится до 6000 на 1 мкм2, и они занимают примерно 70 и 50 % площади тилакоидов гран и тилакоидов стромы соответственно. Фотохимическая активность, электронный транспорт и активность метаболических ферментов ассоциированы с частицами разных типов, и катализируемые ими реакции протекают в направлении, перпендикулярном к мембране. [4]
Структура мембраны и ее фильтрационные свойства определяются, главным образом, параметрами ее формования. Наиболее важными из них являются: продолжительность и температура сушки, температура и состав осадительной ванны, а также толщина слоя раствора. [6]
Исследование структуры мембран показало, что полученные образцы являются сплошными твердыми телами, не обладающими заметной капиллярной пористостью. При взаимодействии с водными растворами возникает вторичная нерегулярная пористость за счет микро - и ультрамикро-трещин, оказывающая влияние на электрохимическую активность таких мембран. [7]
Усадка структуры мембраны, особенно заметная в первые часы работы мембраны, снижает проницаемость и повышает селективность. Спустя сутки после снятия давления характеристики мембраны не восстанавливаются до исходных значений - произошла некоторая остаточная деформация структуры мембраны. В действительности медленное, но непрерывное снижение удельной производительности наблюдается в течение всего срока службы полимерной мембраны. [9]
Нарушение структуры мембраны у мицелиальных грибов под влиянием ПАВ показано в работе ( 21) с помощью метода спиновой метки. [10]
Изучение структуры мембран митохондрий методами рант-гено-структурного анализа и электронной микроскопии позволило сделать заключение, что темные слои стенок мембраны ( рис. 3V) соответствуют - слоям белка, а более светлые - бимолекулярным слоям липоидов. Общая картина строения мембранных стенок митохондрий в настоящее время представляется такой, какой она - показана а рисунке 3V. Каждая мембрана состоит из двух слоев белковых молекул и заключенных между ними двух слоев липидов. На этих белковых и ли-пидных слоях адсорбированы ферменты, которые катализируют биохимические реакции в митохондриях. [11]
На структуру мембран значительное влияние оказывают следующие факторы: тип заряженной частицы, присутствие примесей в полимере, концентрация раствора, вид и длительность дополнительного облучения, частичный отжиг перед выщелачиванием, продолжительность химического травления. При частичном отжиге и применении низкоконцентрированного травильного раствора получаются мембраны с порами малого-диаметра. [12]
На структуру мембран значительное влияние оказывает скорость диффузионных процессов, переводящих систему в области, ограниченные бинодалью или спинодалью, а также скорость собственно разделения на фазы. [13]
Именно сжатием структуры мембраны и выдавливанием из нее части воды, происходящим под действием отрицательного давления, объясняется появление градиента концентрации, показанного на рис. П-49. При малой кривизне менисков ( давление пара под мембраной близко к давлению насыщения) градиент отрицательного давления, а следовательно, и концентрации мал, а при большой кривизне - градиенты велики. С течением времени градиент концентрации сильно меняется, однако это совершенно не сказывается на скорости испарения, которая практически остается постоянной. Его можно просто объяснить, основываясь на предложенном механизме проницания. Вода быстро проницает от одной поверхности к другой по крупным капиллярам, занимающим очень малую часть площади мембраны. Набухание же остальной ее части является медленным процессом и идет с двух сторон мембраны, так как они обе покрыты водой, по направлению к ее средним слоям. Если пренебречь капиллярным характером проницания и воспользоваться полученным распределением концентраций для вычисления коэффициента диффузии, то окажется, что коэффициент диффузии с ростом концентрации уменьшается. Некоторые авторы: этот факт объясняют глобулообразованием. Описанные выше опыты показывают, что его можно объяснить проще, если предположить, что течение через эти мембраны является капиллярным и диффузионный поток в этом случае незначителен. [14]
На податливость структуры мембран могут оказывать влияние некоторые добавки в грунтовку, которые для материала мембраны являются пластификаторами. Так, водные растворы триэтиламина и некоторых спиртов - пластификаторы для ацетатов целлюлозы. Поэтому повышение концентрации триэтиламина или спирта в грунтовке или в промывной воде выше определенной нормы будет ускорять пластическую деформацию мембран и может привести к резкому изменению ее эксплуатационных характеристик. Структурные превращения в мембране под действием давления, очевидно, происходят в макроструктуре и не захватывают тонкую надмолекулярную структуру ацетата целлюлозы. [15]
Страницы: 1 2 3 4
www.ngpedia.ru
Состав мембран
Состав мембран
Основными компонентами мембран являются белки и липиды. На долю углеводов может приходиться около 10% массы мембран, при этом они всегда входят в состав гликолипидов или гликопротеи-нов. Соотношение между белками и липидами в мембранах значительно варьирует — от 20% белка в миелине до 80% в митохондриях. В табл. 1.3 и 1.4 обобщены данные по составу ряда мембран. Плотность мембран прямо пропорциональна содержанию в них белка. Судя по данным изопикнического центрифугирования, чем выше содержание белка в мембране, тем больше ее плотность.
Таблиця 1. Фосфолипидный состав субклеточных мембран печени крысы "
| Доля от суммарного количества фосфолипидов, It | ||||||
| митохондрии | микро-сомы | ли10- сомы | плазматическая мембрана | ядерная мембрана | мембраны аппарата Гольджи | |
| Кардиолипин | 18 | 1 | 1 | 1 | 4 | 1 |
| Фосфатидилэтаноламин | 35 | 22 | 14 | 23 | 13 | 20 |
| Фосфатидилхолин | 40 | 58 | 40 | 39 | 55 | 50 |
| Фосфатидилииозитол | 5 | 10 | 5 | 8 | 10 | 12 |
| Фосфатидилсерин | 1 | 2 | 2 | 9 | 3 | 6 |
| Фосфатидная кислота | — | 1 | 1 | 1 | <1 | |
| Лиэофосфоглицериды 2' | 1 4 | 11 | 7 | 2 | 3 | 3 |
| Сфингомиелин | 1 | 1 | 20 | 16 | 3 | 8 |
| Фосфолипиды | 0,175 | 0,374 | 0,156 | 0,672 | 0,500 | 0,825 |
| Холестерол | 0,003 | 0,014 | 0,038 | 0,128 | 0,038 | 0,078 |
Таблица 2 Белковый и липидиый состав некоторых мембран животных и бактериальных клеток. Л/Б — отношение липка/белок"

Белковый состав мембраны зависит в какой-то степени от метода ее выделения. Некоторые белки непрочно связаны с мембраной и легко удаляются при промывании ее растворами с высокой или, напротив, с низкой ионной силой, щелочными растворами или растворами, содержащими хелатирующие агенты типа ЭДТА. Бывают и случаи, когда трудно сказать, является ли белок мембранным или цитоплазматическим, случайно связавшимся с мембраной в ходе ее выделения.
1. МЕМБРАННЫЕ ЛИПИДЫ
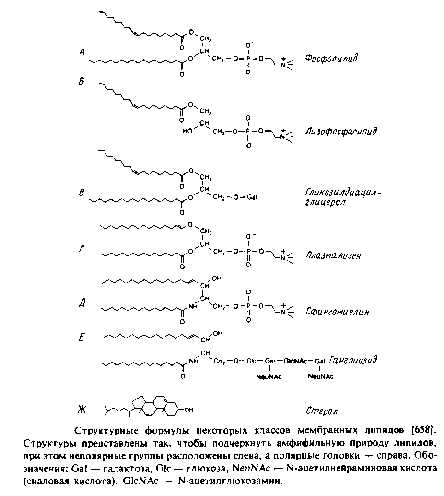
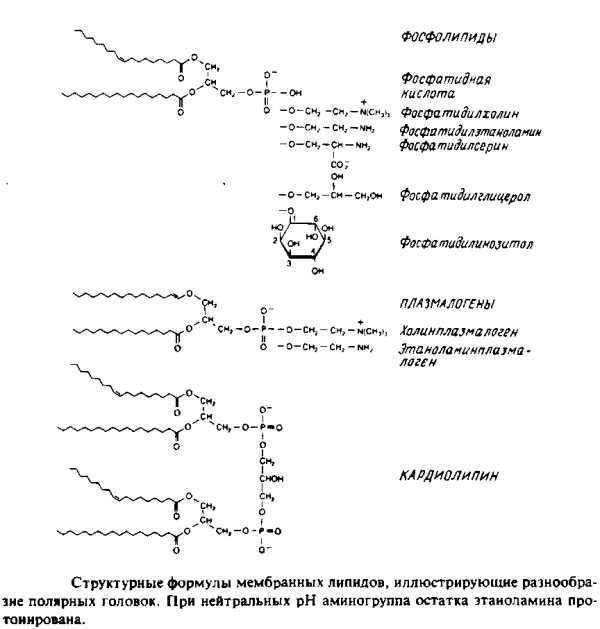
Наиболее поражает в мембранных липидах их огромное разнообразие. Причины этого пока не ясны, хотя становится все более очевидно, что, по-видимому, связано это с тем разнообразием функций, которые липиды выполняют в мембранах. Но, конечно, главная функция мембранных липидов состоит в том, что они формируют бислойный матрикс, с которым взаимодействуют белки. Основные классы липидов представлены на рис. 1.8; их краткое описание дано ниже.
Глицерофосфолипиды
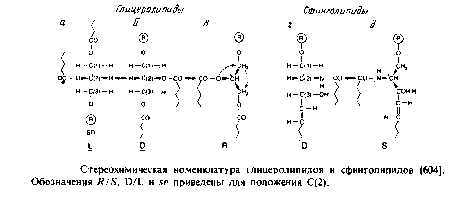
Это наиболее распространенные липиды. Одна из гидроксильных групп глицерола связана с полярной группировкой, содержащей фосфат, а две другие — с гидрофобными остатками. Номенклатура гли-церидов основана на системе стереоспецифической нумерации. Если глицерол изобразить в проекции Фишера, так что центральная группа будет расположена слева, то атомы углерода будут нумероваться так, как показано на рис. В этом случае в название глицерофосфолипида вводят приставку sn для обозначения положения заместителя. В литературе встречается несколько систем стереохимических обозначений: sn, D/L и R/S. На рис. представлена стереохимия атома С-2 в этих трех системах. Природные фосфолипиды.
У большинства фосфоглицеридов фосфатная группа находится в ял-3-положении глицерола; она обычно связана с какой-либо из групп, представленных на рис.


Длинные углеводородные цепи, находящиеся в положениях sn-\ и sn-2, могут присоединяться за счет сложноэфирной и простой эфирной связей. Эти цепи значительно различаются по длине, раз-ветвленности и степени ненасыщенности.
1. 1,2-диацилфосфоглицериды или фосфолипиды. Эти липиды, являющиеся сложными эфирами жирных кислот и глицерола, широко представлены во многих мембранах эукариотических и прокарио-тических клеток, за исключением архебактерий. Фосфатидил-холин является основным компонентом мембран животных клеток, а фосфатидилэтаноламин — это нередко основной липид бактериальных мембран. В табл. представлен ряд жирных кислот, наиболее часто встречающихся в составе фосфолипидов, а в табл. приведен жирнокислотный состав клеточных мембран печени крысы.
Жирные кислоты почти всегда содержат четное число атомов углерода в пределах от 14 до 24. Наиболее распространены кислоты С16, С18 и С20. Степень ненасыщенности может быть разной, но чаще всего встречаются ненасыщенные кислоты 18:1, 18:2, 18:3 и 20:4. Здесь первое число обозначает длину ацильной цепи, а второе указывает на число содержащихся в ней двойных связей. Почти все природные кислоты характеризуются г/ис-конфигурацией двойных связей. Цепь в такой конфигурации имеет излом, что нарушает упаковку липидных молекул в бислое. В составе молекул многих фосфолипидов имеется одна насыщенная и одна ненасыщенная цепи. В случае животных клеток ненасыщенные цепи обычно находятся в 5и-2-положении глицерола. Такое же распределение цепей характерно и для фосфолипидов клеток Е. coli. Двойные связи в полиненасыщенных цепях обычно являются несопряженными. В фосфолипидах некоторых бактериальных мембран обнаружены разветвленные цепи, а также цепи, содержащие циклы, и гидроксильные группы в /3-положении. На рис. 1.11 показана структура некоторых из этих кислот.
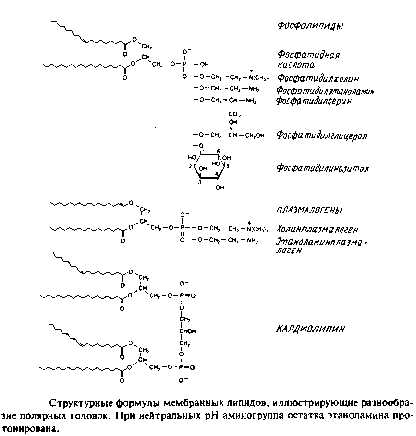
У архебактерий глицерофосфолипиды имеют обращенную сте-реохимическую конфигурацию, при которой фосфорильные группы находятся в sn-\-положении глицерола. У многих бактерий этого вида гидрофобные компоненты представляют собой не сложные эфиры жирных кислот, а простые изопранильные эфиры глицерола.
Кардиолипины или дифосфатидилглицеролы. По сути это димерные формы фосфолипидов. Они содержатся в боль шом количестве во внутренней мембране митохондрий, в мембране хлоропластов и в некоторых бактериальных мембранах, но редко встречаются в других мембранах.

4. Плазмалогены. Это фосфоглицеролипиды, у которых одна из углеводородных цепей представляет собой простой виниловый эфир. Этаноламиновые плазмалогены широко представлены в миелине и в саркоплазматическом ретикулуме сердца.
Фосфосфинголипиды
Эти липиды имеют такие же полярные головки, как и глицерофосфолипиды, но их гидрофобная часть представлена церамидом. В плазматических мембранах животных клеток широко распространен сфингомиелин. Основными жирнокислотными компонентами в миелине являются кислоты 24:1 и 24:0. В мембранах растительных и бактериальных клеток фосфосфинголипиды встречаются редко. Кроме сфингомиелина известны и другие фосфосфинголипиды, например церамид-1-фосфорилэтаноламин, церамид-1-фосфорилинози-тол и церамид-1-фосфорилглицерол.
Гликоглицеролипиды
Это полярные липиды, у которых в м-З-положении глицерола находится углевод, присоединенный с помощью гликозидной связи, например галактоза. Гликоглицеролипиды широко представлены в мембранах хлоропластов, они обнаружены также в заметных количествах в сине-зеленых водорослях и бактериях. Моногалактозилдиа-цилглицерол был назван «наиболее распространенным в природе полярным липидом», поскольку на его долю приходится половина всех липидов тилакоидной мембраны хлоропластов. Для мембран грамположительных бактерий характерны гликоглице-
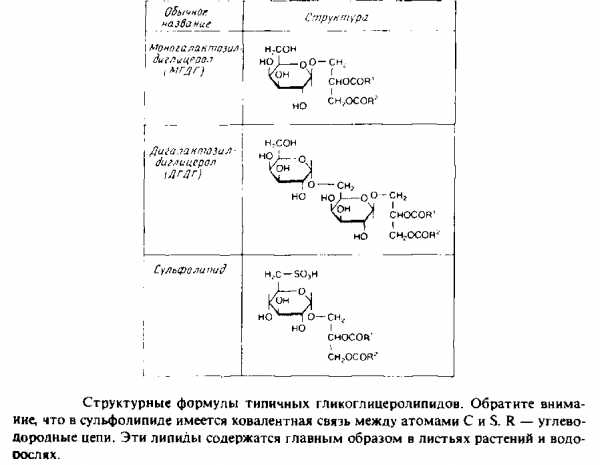
ролипиды с большим разнообразием Сахаров. Архебактерии также содержат такие липиды, но, как и в случае глицерофосфолипидов, их стереохимическая конфигурация является обращенной, с локализацией гликозидной связи в sn-l-положении глицерола. В мембранах животных клеток гликоглицеролипиды встречаются редко.

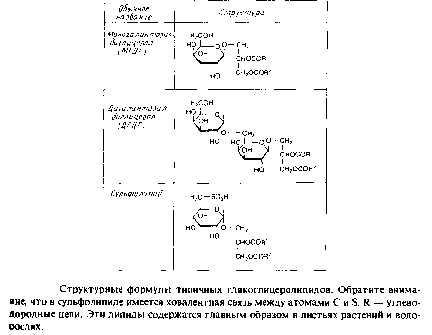
Гликосфинголипиды
классифицируют в соответствии с размером углеводной части, которая может быть представлена всего лишь одним моносахаридным остатком, с одной стороны, и очень сложным углеводным полимером — с другой. Моногликозилцерамиды обычно называют цереброзидами. Ганглиозиды представляют собой класс анионных гликосфинголипидов, которые содержат один или несколько остатков сиаловой кислоты, связанных с сахарными остатками церамидолигосахарида. Глобозидами называют нейтральные гликосфинголипиды, которые не содержат остатков отрицательно заряженной сиаловой кислоты.
Гликосфинголипиды находятся на наружной поверхности плазматических мембран животных клеток; обычно они являются минорными компонентами, но иногда содержатся в значительных количествах. Моногалактозилцерамид — это один из основных компонентов миелиновой оболочки нервного волокна. В некоторых случаях гликосфинголипиды локализуются не в плазматической мембране, а во внутриклеточных мембранах.
Гликосфинголипиды мембран эритроцитов несут антигены группы крови. В клетках аденокарциномы человека накапливаются необычные фукозилированные гликосфинголипиды, которые можно использовать для обнаружения этих клеток и контроля за развитием опухоли.
Стеролы
Эти липиды присутствуют во многих мембранах растений, животных и микробов. По-видимому, самым распространенным из сте-ролов является холестерол. Его молекула состоит из компактного, жесткого гидрофобного ядра, а полярной головкой является гидро-ксильная группа. Холестерол содержится в плазматических мембранах животных клеток, в лизосомах, эндосомах и в мембранах аппарата Гольджи. Он составляет около 30% всей массы мембранных липидов во многих плазматических мембранах животных клеток. В высших растениях обнаружены другие стеролы, чаще всего ситостерол и стигмастерол. Растительные стеролы часто имеют еще одну боковую цепь в положении С-24 и/или двойную связь в положении С-22. В мембранах дрожжей и дру-
гих эукариотических микроорганизмов часто содержится эргостерол. К классу стеролоподобных липидов относят также гопанои-ды, которые найдены в бактериях и некоторых растениях.
Минорные компоненты
В мембранах присутствуют также и другие липиды, которые можно отнести к разряду минорных компонентов вследствие их малого содержания в мембранах. Так, в мембранах обычно обнаруживаются, хотя и в очень малых количествах, свободные жирные кислоты и лизофосфолипиды. Пожалуй, исключением из этого правила являются мембраны хромаффинных гранул, которые, как известно, содержат необычно много свободных жирных кислот. Минорными компонентами мембран являются также моноацил- и диацилглице-ролы. Диацилглицеролы выполняют важную функцию вторых посредников в передаче сигнала при активации клеток рядом биологически активных веществ. Эта система клеточного отклика на внешний стимул будет детально рассмотрена в гл. 9. В мембранах обычно присутствуют и полиизопреноидные липиды. К ним относятся уби-хиноны и менахиноны — компоненты цепи электронного транспорта в мембранах. Можно отметить также ундекапренол и долихол, которые являются липидными переносчиками промежуточных продуктов соответственно при биосинтезе клеточной стенки у прокариот и при биосинтезе гликопротеинов в аппарате Гольджи эукариот. Длина молекул этих липидов в вытянутом состоянии значительно превышает толщину бислоя, поэтому неизвестно, как эти молекулы в нем расположены. Неясно также, почему липидными переносчиками служат именно полиизопреноидные структуры.
2. МНОГООБРАЗИЕ ФУНКЦИЙ МЕМБРАННЫХ ЛИПИДОВ
Совершенно очевидно, что липидный состав различных мембран не является случайным, однако удовлетворительного объяснения этому феномену не найдено. Любая конкретная мембрана может содержать более ста разных типов липидных молекул. Почему их так много и почему каждая мембрана имеет уникальный липидный состав? Пути биосинтеза мембранных липидов и механизмы их доставки к местам назначения обсуждаются в разд. 10.4. Становится все более очевидным, что липиды активно участвуют в процессах, протекающих в мембранах, однако причины их разнообразия неясны. Рассмотрим некоторые факторы, возможно, определяющие липидный состав мембраны.
1. Смесь липидов обязательно должна быть способна образовать стабильный бислой, в котором могли бы функционировать белки.
Некоторые липиды способствуют стабилизации сильно искривленных участков мембраны, образованию контакта между мембранами или связыванию определенных белков, поскольку форма этих молекул благоприятствует нужной упаковке бислоя на соответствующих участках мембраны.
Некоторые липиды являются важными биорегуляторами. Наиболее изучена в этом отношении регуляторная роль производных фосфатидилинозитола в плазматических мембранах клеток эукариот.
Некоторые липиды участвуют в реакциях биосинтеза. Например, в клетках Е. coli фосфатидилглицерол поставляет глицерофосфатный фрагмент при биосинтезе периплазматических олигосахари-дов.
Отдельные липиды необходимы для поддержания оптимальной активности ряда ферментов. Этот вопрос рассматривается в гл. 6.
Ганглиозиды, как полагают, играют важную роль в регуляции роста клеток, являются специфическими рецепторами в плазматической мембране и ответственны за клеточную адгезию.
Как было показано экспериментально, организмы часто могут выдерживать — причем без всяких последствий — существенные изменения липидного состава мембран. Например, с помощью генетической трансформации можно получить штаммы Е. coli, в мембранах которых содержится 34% фосфатидной кислоты, обычно отсутствующей в штаммах дикого типа. Очевидно, тот липидный состав, который характерен для штаммов дикого типа, не является обязательным для выживания клеток, по крайней мере, в условиях их выращивания в лаборатории.
3. МЕМБРАННЫЕ БЕЛКИ
Как видно из табл. 1.3 и 1.4, мембраны содержат от 20 до 80% белка. Как правило, именно белки ответственны за функциональную активность мембран. К ним относятся разнообразные ферменты, транспортные белки, рецепторы, каналы, поры и т. д., которые обеспечивают уникальность функций каждой мембраны. Первые успехи в изучении мембранных белков были достигнуты тогда, когда биохимики научились использовать детергенты для выделения мембранных белков в функционально активной форме. Это были работы по изучению ферментных комплексов внутренней мембраны митохондрий. Значительным шагом вперед было осознание того, что мембранные белки имеют не исключительно /3-складчатую структуру, как предполагалось в модели «элементарной мембраны» Дэвсона—Даниелли—Робертсона, а содержат достаточно много а-спиралей. Важное значение имел также вывод о том, что мембранные белки могут глубоко проникать в липидный бислой или даже пронизывать его и что их стабилизация осуществляется за счет гидрофобных взаимодействий. Эти термодинамические представления существенно обогатили принцип «гидрофобных сил», предложенный для объяснения структуры белков и предполагавший существование неполярной, гидрофобной области внутри белковой глобулы и полярных, гидрофильных участков, контактирующих с водной средой.
По мере совершенствования методов очистки удавалось получать в изолированном виде все большее число мембранных белков. Определение первичной структуры большинства из них было затруднено из-за плохой растворимости в воде как самих белков, так и получаемых из них гидрофобных пептидов. В середине 1970-х гг. эта проблема была решена для двух мембранных белков — гликофорина и цитохрома bs, что позволило установить основной принцип структурной организации интегральных белков. В аминокислотной последовательности гликофорина — сиалогликопротеина из мембраны эритроцитов — был обнаружен короткий участок, состоящий из 23 неполярных аминокислот и расположенный примерно в середине цепи. Данные топологических и других исследований показали, что молекула гликофорина полностью пронизывает мембрану, причем погруженный в мембрану гидрофобный участок имеет а-спиральную конфигурацию. Так вошла в жизнь новая, теперь уже общепризнанная концепция о наличии в мембранных белках а-спиральных доменов, пронизывающих мембрану. Эта концепция была полностью подтверждена при изучении трансмембранных белков с помощью методов, которые позволяют получить максимально возможное в наше время разрешение. Судя по результатам реконструкции электронно-микроскопических изображений препаратов бактериородопсина из пурпурной мембраны Halobacterium halobium и по данным рентгеноструктурного исследования фотосинтетических реакционных центров бактерий, эти белки содержат несколько а-спиральных участков, последовательно пересекающих бнслой.
Другой вариант расположения полипептидной цепи в мембране был обнаружен при изучении аминокислотной последовательности интактной формы микросомного цитохрома bi. Было показано, что этот белок содержит относительно короткий участок вблизи карбоксильного конца, состоящий из гидрофобных аминокислот. Этот «гидрофобный якорь» можно было удалить с помощью протеолиза, причем гемсвязывающий домен высвобождался в водорастворимой форме. Локализованный в мембране гидрофобный домен, или «якорь», стал еще одним характерным элементом структуры мембранных белков.
В основе современных представлений о структуре мембранных белков лежит идея о том, что их полипептидная цепь уложена так, чтобы образовалась неполярная, гидрофобная поверхность, контактирующая с неполярной областью липидного бислоя. Полярные или заряженные домены белковой молекулы могут вазимодействовать с полярными головками липидов на поверхности бислоя. Многие мембранные белки являются трансмембранными и пронизывают бислой. Некоторые белки, по-видимому, связаны с мембраной лишь за счет их взаимодействия с другими белками.
Мембранные белки обычно связываются с мембраной с помощью нековалентных взаимодействий — гидрофобных или электростатических сил. Однако есть мембранные белки, которые связаны с липидами ковалентно. Такие примеры пока немногочисленны, но их появляется все больше. Многие белки плазматических мембран растительных и животных клеток относятся к классу гликопротеинов. Углеводные остатки этих белков всегда находятся с наружной стороны плазматической мембраны.
Обычно мембранные белки подразделяют на наружные и внутренние. При этом критерием служит степень жесткости обработки, необходимой для извлечения этих белков из мембраны. Периферические белки высвобождаются при промывании мембран буферными растворами с низкой ионной силой, буферными растворами с низким или, наоборот, высоким значением рН и в присутствии хелатирующих агентов, связывающих двухвалентные катионы. Как полагают, такие белки связаны с поверхностью мембраны за счет слабых электростатических взаимодействий с полярными головками липидных молекул либо с молекулами других белков. Часто бывает нелегко отличить периферические мембранные белки от белков, связавшихся с мембраной в процессе ее выделения. При обработке мембранного препарата буфером с низкой ионной силой в раствор переходит около 30% белков, связанных с мембраной эритроцитов. При несколько более жесткой обработке хаотропными агентами высвобождаются периферические белки. В ряде случаев эти агенты оказывают влияние достаточно сильное, чтобы разрушить белок-белковые взаимодействия, хотя денатурации белков при этом не происходит. Действие хаотропных агентов, «нарушающих структуру воды», обусловлено главным образом ослаблением гидрофобных взаимодействий между компонентами мембраны.
Для высвобождения интегральных мембранных белков необходимо использовать детергенты или даже органические растворители. Детергенты разрушают липидный бислой и, как полагают, связываются с гидрофобными участками мембранных белков, контактирующими с гидрофобной областью бислоя. Для того чтобы сохранить интегральные мембранные белки в растворенном монодисперсном состоянии, в растворе постоянно должны присутствовать детергенты. При удалении детергентов неизбежно происходят агрегация белков и их последующее осаждение. Дальнейшая информация о взаимодействиях между белками и детергентами, а также о структуре и свойствах мембранных белков содержится в гл. 3.
Резюме
Методы дифракции рентгеновских лучей и электронной микроскопии сыграли историческую роль в развитии мембранологии и внесли решающий вклад в современные представления о биологических мембранах. Сегодня не вызывает сомнений, что липидный бислой образует структурную основу практически всех биологических мембран и что их функциональное многообразие основано именно на этом структурном единстве различных мембранных систем. Каждая отдельно взятая мембрана содержит большое число различающихся по своим химическим свойствам липидов. Причины этого разнообразия неясны, хотя появляется все больше данных об уникальных биологических функциях отдельных липидов. На долю белков приходится от 20 до 80% массы мембран. Многие из этих белков полностью пронизывают липидный бислой, и их удается солюбилизировать только с помощью детергентов. Другие мембранные белки, называемые периферическими, легкое извлекаются из мембраны с помощью буферных растворов с изменяющимися рН или ионной силой либо при удалении двухвалентных катионов с помощью хелатирующих агентов.
В настоящее время разработаны методы выделения и характеристики индивидуальных мембран из клеток прокариот, из животных и в меньшей степени из растительных клеток. Разделение чаще всего основано на различиях в размере и плотности мембранных частиц, содержащихся в гомогенате разрушенных клеток. Можно также использоать различия в поверхностных свойствах мембран или в их электрофоретическом поведении. Выделение и очистка мембран — первый и обязательный этап в их биохимическом исследовании.
baza-referat.ru